



Número actual.

Abril-Junio 2026 vol. 77-2
Portada
Indice
Los cactus: modelos únicos para la ciencia
Los viajes al espacio, un reto para el cuerpo humano
¿Cómo influye tu microbiota en la ansiedad?
Acciones para el control de bacterias resistentes a los antibióticos
Metástasis: células cancerosas que no respetan fronteras
Los microARN, reguladores y controladores de funciones biológicas
El misterioso mundo de los glicoARN
Cómete los tacos, ¡pero con salsa!
El mundo nanoscópico de los virus a través de la luz
Armonía a escala nanométrica
La memoria episódica es más que un cúmulo de recuerdos
¿Todo es medible? La métrica de las cosas
Evaluación de la calidad del tequila
Aprovechamiento de efluentes vinícolas para generar nuevos productos
El desarrollo orientado al transporte (DOT) y su aplicación en México
Tales de Mileto: uno de los siete sabios de Grecia
De moléculas a megafábricas. ¿Cómo se diseñan las transformaciones químicas que mueven el mundo?
Desde las redes
Noticias de la AMC
Desde el Comité Editorial
Artículos del número anterior.
Indice
Noticias de la Academia Mexicana de Ciencias
Portada
La degradación ambiental de la Cuenca del Alto Atoyac y su impacto en la salud
Marcadores moleculares del ADN: huellas genéticas que revelan secretos de los seres vivos
Microorganismos que ayudan a descontaminar agua con arsénico
Mitos comunes sobre la anticoncepción tras el embarazo adolescente
Las barreras que dificultan una buena lactancia en madres adolescentes
La importancia de la salud bucodental durante el embarazo de las adolescentes
Alimentación y hábitos saludables en las adolescentes embarazadas
La programación de la vida desde el vientre: nutrición, genética y epigenética
Hormonas, metabolismo y bioquímica durante el embarazo de una adolescente
Transformación de la identidad en el embarazo adolescente
La búsqueda de afecto y familia en el contexto del embarazo adolescente
El papel de la dinámica familiar en el embarazo adolescente
El embarazo adolescente es un asunto de la familia, la escuela y el Estado
Embarazo adolescente, una visión integral
Desde el Comité Editorial
vol. 58 num. 4 octubre - diciembre 2007

Beneficios de los sistemas agroforestales: amor por nuestras tierras
Manuela Tamayo-Chim y Roger Orellana
![]()
![]()
| El c o n t r o l | |||
| del ciclo celular | |||
| en plantas | |||
| Juan G. Colli-Mull, Luis Carlos Rodríguez Zapata, Enrique Castaño de la Serna |
|||
|
|
|||
|
Introducción |
|||
|
|
|||
| Figura 1. Organización estructural y propiedades de la familia de E2F en Arabidopsis. En la familia de E2F se conservan los dominios de unión al ADN, de dimerización, de caja de marcaje y el dominio de transactivación. Basado en la conservación de los diferentes dominios, las ocho proteínas de E2F de Arabidopsis son clasificadas en E2F, DP y DEL. |
|||
|
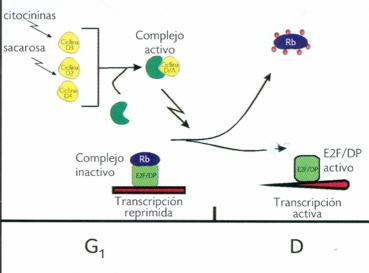
Una ruta análoga de E2F/pRb en la transición G1-S del ciclo celular, que involucra la fosforilación de pRb por los complejos CDK/ciclina y la liberación de E2F, ha sido dilucidada recientemente en células vegetales, y probablemente los mecanismos de su regulación sean los mismos que en sus contrapartes en humanos y otros eucariontes superiores (Figura 2). Aunque no se han encontrado homólogos de las ciclinas E en plantas, fundamentales en la transición G1-S en animales, las ciclinas tipo A o quizá algunas del tipo D podrían participar en la regulación de la transición G1-S y de G2-M del ciclo celular (Wen-Hui Sen, 2001). |
|||
|
Figura 2. Modelo propuesto para la regulación de la transición G1-S del ciclo celular en plantas. La expresión de las ciclinas D es estimulada por señales del medio ambiente y de crecimiento. Estas ciclinas se pueden asociar a la CDK-A. Este complejo activo ciclina D/CDK-A puede fosforilar a la Rb, la cual se encuentra formando un complejo con E2F/DP, inhibiendo la transcripción. Conforme avanza el ciclo celular, Rb es hiperfosforilada por el complejo ciclina D/CDK-A en G1 tardía, lo que ocasiona que se libere el complejo transcripcional E2F/DP, el cual es capaz de activar la transcripción de genes requeridos para la fase S del ciclo celular. |
|||
|
Genes regulados por E2f que participan en el ciclo celular |
|||
|
|
|||
|
|
|||
|
Juan Gualberto Colli-Mull es licenciado en Biología por el Instituto Tecnológico Agropecuario No. 2, y maestro en Ciencias y Biotecnología de plantas por el Centro de Investigación Científica de Yucatán (cicy). Actualmente está cursando el doctorado en Ciencias y Biotecnología de Plantas en el CICY. Su trabajo se centra en el estudio de la función de E2F en procesos de desarrollo y apoptosis en cultivos in vitro de Coffea canephora. Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo. |
|||
|
Bibliografía |
|||


| ¿Para qué sirve | |
|
la endogamia?
Fidel Márquez Sánchez
|
|
|
La endogamia, que el diccionario define como el “cruzamiento entre individuos de una raza, comunidad o población aislada genéticamente”, se caracteriza técnicamente como la condición homocigótica de genes en un determinado sitio (locus) cromosómico. |
|
|
|
Lo malo de la endogamia El efecto perjudicial de la endogamia, también llamado “depresión endogámica”, necesariamente tuvo que ser observado por el ser humano, probablemente dentro del mismo grupo al que pertenecía, al descubrir que los hijos (progenie) de parejas emparentadas mostraban anomalías como enanismo, albinismo, hemofilia, etcétera, que se acentuaban a lo largo de las generaciones. Para evitarla debió comenzar a hacer precisamente lo contrario del apareamiento endogámico, es decir, procurar que los cruzamientos se hicieran entre individuos no emparentados. |
|
Al reconocer el efecto de la endogamia en las poblaciones de animales o plantas que explotaba, el ser humano comenzó a “refrescar la sangre” mediante apareamientos con miembros de otras poblaciones no emparentadas y diferentes a las suyas. En las ganaderías empresariales se procura conocer el pedigrí de los pies de cría; es decir, se hace una selección de los progenitores. De hecho, podemos decir que las formas para evitar la endogamia (o “consanguinidad”, como prefieren llamarla los zootecnistas y ganaderos) tuvieron lugar primero en la cría y explotación del ganado, ya que en los animales sus efectos son más evidentes que en los cultivos agrícolas. |
|
|
|
|
|
De que los genes en un locus de un individuo sean idénticos a los genes de un locus de otro individuo. Por lo tanto, la coancestría entre dos progenitores es igual a la endogamia de su progenie. De hecho, en la práctica, la endogamia de una progenie es igual a la coancestría de sus progenitores. Por ejemplo, si X y Y son los progenitores de Z, entonces rXY = FZ |
|
|
|
|
|
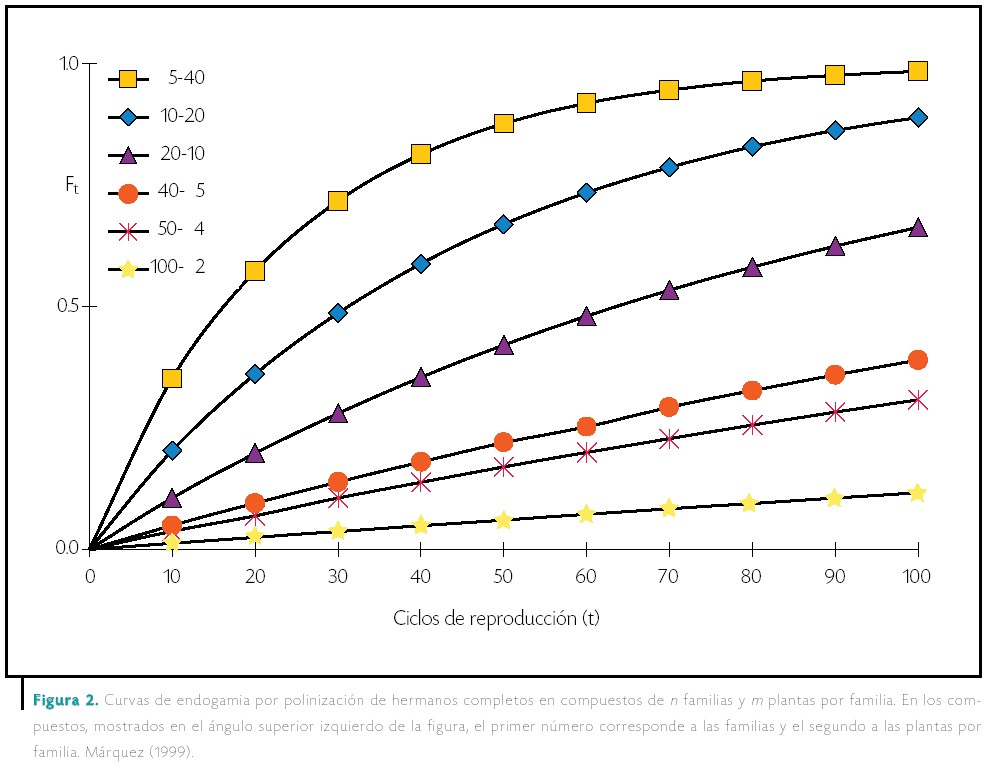
Los resultados de un estudio de Márquez (1992) sobre la endogamia en los sintéticos mostraron que mientras mayor era el número de líneas (de 2 a 1) la reducción de la endogamia era de casi 0 a 80 por ciento para cualquier grado de endogamia de las líneas, mientras que si se aumentaba el número de plantas por línea (de 1 a 10), las reducciones eran de 90 por ciento para plantas no endogámicas, 45 por ciento para líneas fraternales, 50 por ciento para líneas S1 y 0 por ciento para líneas homocigóticas. |
|
|
|
|
|
Incremento del germoplasma de maíz |
|
 |
|
|
Entonces, ¿para qué sirve la endogamia? |
|
| Bibliografía Busbice, T. H. (1969), "Inbreeding in synthetic varieties", Crop Sci. 9:601-604. Busbice, T. H. (1970), "Predicting yield of synthetic varieties", Crop Sci. 10:260-269. Falconer, D. S. (1967), Introduction to quantitative genetics, Oliver and Boyd, Londres, pp.85-93. Márquez Sánchez, F. (1988), Genotecnia vegetal II, AGT Editor, México, 665 p. Márquez-Sánchez, F. (1992), "Inbreeding and yield prediction in synthetic varieties of maize cultivars made with parental lines: I. Basic methods", Crop Sci. 32:345-349. Márquez-Sánchez, F. (1998), "Expected inbreeding with recurrent selection in maize: I. Mass selection and modified ear-to-row selection", Crop Sci. 38:1432-1436. Márquez-Sánchez, F. (1999), "Accumulated inbreeding in maize germplasm reproduction", Maydica 44:225-229. Márquez-Sánchez, F. (2005), "Derivation of open-pollinated inbred lines, and their relation to Z lines for cyclic hybridización", Arnel R. Hallauer international symposium in plant breeding, CIMMYT, agosto17-22 (acepta-do para publicación). Wright, S. (1921), "Systems of mating II. The effects of inbreeding on a genetic composition of a population", Genetics 6:124-143. |
|
| Fidel Márquez Sánchez es ingeniero agrónomo egresado de la Escuela Nacional de Agricultura en 1958. Obtuvo la maestría y el doctorado en la Iowa State University. Es doctorado honoris causa y agrónomo ilustre de la Universidad Autónoma de Chapingo, y fellow de la American Society of Agronomy. Pertenece a la Academia Mexicana de Ciencias y al Sistema Nacional de Investigadores. Desde hace 14 años trabaja en el Centro Regional Universitario de Occidente de la Universidad Autónoma de Chapingo, en Guadalajara, Jalisco, en el campo de mejoramiento genético de maíces criollos e hibridación. Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo. | |



y los
![]()
![]()
Regreso al origen
Vicente Aboites![]()
![]()
|
Noticias de la |
|||
| Academia | |||
| Olimpiadas de la Ciencia | |||
| Triunfa México en competencias internacionales de ciencias | |||
|
Este año la Academia Mexicana de Ciencias obtuvo varias medallas en competencias internacionales, entre ellas la medalla de oro en el Octavo Campeonato Mundial de National Geographic, cuya final se celebró el pasado 9 de agosto, en el SeaWorld de San Diego, California, dejando al equipo de Estados Unidos -el anterior campeón- en el segundo sitio y a Canadá en tercero. |
|||
|
Carlos Bosch Giral, coordinador de la Competencia Cotorra y del Concurso de Primavera de Matemáticas, mencionó que estos resultados son sólo la punta del iceberg del esfuerzo conjunto y del trabajo en equipo que involucra a la AMC, a profesores y a familiares, y que ahora rinde sus frutos con medallas internacionales. |
 |
||
| Los adolescentes y jóvenes triunfadores, junto al Dr. Juan Pedro Laclette, de visita en las instalaciones de la AMC. | |||
|
Consejo directivo |
|||
| Crean fondo para apoyar a jóvenes con talento científico | |||
|
El presidente de México, Felipe Calderón, anunció la creación de un fondo de becas para apoyar a los ganadores de las olimpiadas científicas con el objetivo de impulsar su talento, que será coordinado por la Academia Mexicana de Ciencias (AMC) y la Secretaría de Educación Pública (SEP). Al respecto, Juan Pedro Laclette, presidente de la AMC, subrayó que la organización de las olimpiadas, junto con la detección de jóvenes |
|||
|
|
Felipe Calderón anunció la creación del fondo, durante la visita de los ganadores del Octavo Campeonato de National Geographic a Los Pinos. |
||
|
Novedad editorial |
|||
| Rinden homenaje a pioneras de la ciencia en México | |||
| La Academia Mexicana de Ciencias (AMC), el Consejo Consultivo de Ciencias de la Presidencia de la República y el Consejo Nacional de Ciencia y Tecnología rinden homenaje a mujeres científicas con el quinto volumen de la serie Ciencia y tecnología en México en el siglo XX. Presentado en junio pasado, el libro Biografías de personajes ilustres, dedicado a mujeres forjadoras de la ciencia y la tecnología mexicanas, describe las carreras de 14 investigadoras mexicanas, cinco de ellas dedicadas a las ciencias naturales y exactas y otras nueve enfocadas al ámbito de las ciencias sociales y humanidades. Juan Pedro Laclette, presidente de la AMC, mencionó que, si bien a mediados del siglo pasado la mujer se integró al campo laboral y se han reducido las diferencias entre ambos sexos, aún hay mucho que hacer. Por ello, la Academia mantiene un enfoque de género en todo momento. "La AMC cuenta entre sus integrantes con un mayor porcentaje de mujeres que otras academias del primer mundo, con un 21 por ciento", afirmó Laclette. "En academias de ciencias europeas como las de Francia, Alemania y Dinamarca, entre otras, el porcentaje de mujeres es mucho más bajo". |
|||
|
|
|||
|
El quinto volumen de la serie Ciencia y Tecnología |
|||


|
ciencia |
|
|
octubre-diciembre 2007 volumen 58 número 4 |
|
|
comunicaciones libres |
|
Reflexiones sobre la investigación en arquitectura Fernando N. Winfield Reyes La precisión en los cálculos científicos por computadora Edscott Wilson García y Guillermo Morales-Luna Pinzas ópticas: las delicadas manos de la luz Karen Volke Sepúlveda, Ibis Ricárdez Vargas y Rubén Ramos García La codificación de los quipus incas Hugo Cabrera Ibarra, Haret Rosu Barbus, Luís Adolfo Torres González y Juan Pablo Treviño Gutiérrez El desierto y sus habitantes: el caso de la hormiga roja Rafael Guzmán Mendoza Las selvas tropicales y el cambio climático Julián Granados Cambio de uso del suelo y degradación ambiental Leopoldo Galicia Sarmiento, Arturo García Romero, Leticia Gómez-Mendoza y M. Isabel Ramírez Selva seca y sequía: compañeras inseparables Mirna Valdez-Hernández, Elizabeth Ma. Osorio-Gil, Paula Jackson y José Luís Andrade |
editorial Juan Pedro Laclette
Beneficios de los sistemas agroforestales: amor por nuestras tierras El control del ciclo celular en plantas ¿Para qué sirve la endogamia? Los tribunales constitucionales y la división de poderes debate |
